I've recently been struggling with this very question, and think I have figured it out for myself. Here's the way I came to understand it:
Couple notes:
First, much of this insight is due to Kalid Azad and Grant Sanderson.
Second, I'm describing exponential growth as a function of time here, but it could easily be a function of anything else, such as distance, etc.
Suppose you have a single individual creature, with mass $1$ kg of "active tissue", at day $0$. The creature grows in the following way: At the start of each day, the active tissue spawns new, inactive tissue, at a linear rate such that at the end of that day, the amount of new tissue is equal to that of the starting (active) tissue. And at the moment this doubling has been completed, the newly spawned tissue also becomes active.
Such a function would look like this:
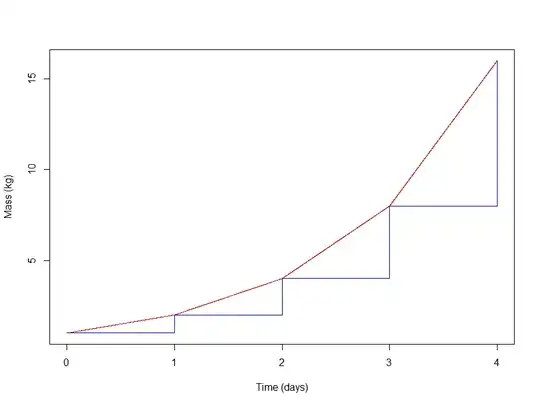
The red line shows the mass as a function of time, and the height of each blue horizontal line indicates the mass of the active tissue during that particular day. Note that the function is noncontinuous.
Next, suppose we want to consider the rate at which this function grows. There are two distinct things we could mean by "rate".
1) The relative growth rate, which is the rate the active tissue grows, relative to the mass of the active tissue. At the start of day $0$, there is $1$ kg of active tissue. At the end of day $0$, there is $1$ kg of new tissue. So the relative growth rate is ($1$ kg new tissue/$1$ kg active tissue)/$1$ day, which is $1$ unit/day, or $100\%$ growth per day.
At the start of day $1$, the newly formed tissue becomes active, so now we have $2$ kg of active tissue. At the end of day $1$, we have $2$ kg of new tissue. So the growth rate is ($2$ kg/$2$ kg)/$1$ day = $100\%$ growth/day.
What if we restrict our time period to half a day? Let's choose the start of day $1$ as our start time, and halfway through day $1$ as our end time. During this half day, we've grown $1$ kg of new tissue, and this was born of $2$ kg of active tissue. So the growth rate is ($1$ kg/$2$ kg)/$0.5$ days = $100\%$ growth/day. So, despite this strange noncontinuous function, the growth rate is a constant $100\%$ per day.
2) The time derivative of mass, $\frac{dm}{dt}$, which denotes how fast the mass is changing per unit time. We're now not asking how much it's changing proportionally, but how much the mass variable itself is changing. As such, it will be in kg/day, rather than units/day or percentage growth/day.
Notice that at any given time, $\frac{dm}{dt}$ is equal to the current amount of active tissue (i.e. the total accumulated mass at the start of that day). And this makes perfect sense - if the active tissue is growing at $100\%$ per day, then $\frac{dm}{dt}$, at any given moment, is $100\%$ of the mass of the currently active tissue. You can verify this by inspecting the graph: The height of the horizontal blue lines (position on the y-axis that they'd intersect if extended) indicates two things: the mass of active tissue during that day, and also $\frac{dm}{dt}$, or the slope, during that day. Those values are $1,2,4,8$ for the four days shown.
Also note that, unlike $\frac{dm}{dt}$, the relative growth rate is invariant. It is always $100\%$ no matter what the time is. As such, it is a particularly useful quantity, since it efficiently captures much of the information about this function. Keep this fact in mind, as it will be relevant later, when addressing your specific question.
Now, let's modify the system in a crucial way. Instead of having to wait a whole day before the new tissue becomes active, new tissue is immediately active from the moment it is born. We now have continuous $100\%$ growth/day. As a result, the total tissue at the end of the first day will be more than $1$. It will be $e$. This is the result of deriving $e$ from continuously compounding interest, as demonstrated in user223391's answer.
So $e$ is the ratio of how much you end up with to how much you started with, when you have continuous growth at a rate of $100\%$ per day, for a duration of $1$ day.
So, if under continuously compounding growth, we end up with $e$ kg at the end of the first day (starting with $1$ kg), what will be our total mass at the end of day $2$? Well, remember that we defined $e$ as the ratio of how much you end up with to how much you started with. If we start with $e$, then at the end of another unit of time, we'll have $e\cdot e$, or $e^2$. In general, the growth can be modeled by
$$m=e^t$$
which represents mass as a function of time, under a regime of continuous compounding $100$ percent growth.
Side note: technically, a function can be continuous, but not continuously compounding. $y = x$ and $y = x^2$ are both continuous, but they are not continuously compounding. Any function with the form $A \cdot B^X$ is, however, continuously compounding. It's probably safe to assume that anytime you see the term "continuous growth", it means "continuously compounding growth".
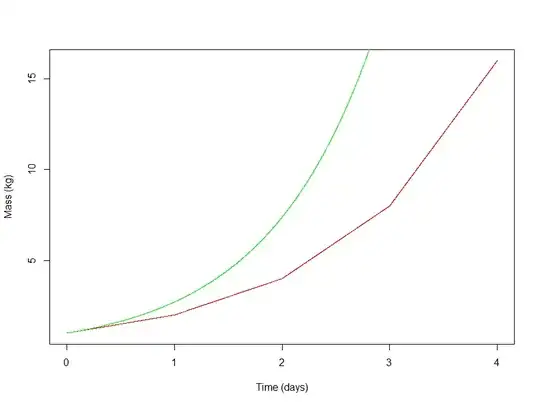
Notice that this new function (green) rises faster than the noncontinuous (red) function. This isn't surprising, given that $e$ is larger than $2$.
What about $\frac{dm}{dt}$ with this new system? Well, that's easy. It's the same as before we had continuous growth. $\frac{dm}{dt}$ is equal to the amount of active tissue that currently exists, per unit time (kg/day). Except that now, all the tissue is active! So, $\frac{dm}{dt}$ is now a continuous function, and evolves as gradually as the amount of active mass does, which is infinitely gradually! Compare this to the noncontinuous function, where $\frac{dm}{dt}$ is still equal to the amount of active tissue, but since the amount of active tissue grows discretely, so does the time derivative. Also note that in the noncontinuous case, $\frac{dm}{dt}$ is not equal to the total amount of accumulated tissue, as it is in the continuous case, since not all of the accumulated tissue is active.
So we've talked about $100\%$ growth (from here on in, "growth" means "continuous growth"). What about $50\%$ growth? How would we model that using $e$?
Well, let's work backwards. First, let's assume we can model this with $e^x$, where $x$ is a function of $t$, which we'll call $g(t)$. Somehow, $g(t)$ incorporates information about our growth rate. Under this assumption, when the growth rate is $1$, $g(t)$ must be equal to $t$. So, our more general model of growth, that isn't restricted to $100\%$ growth, is
$$m = e^{g(t)}$$
At 50% growth per day, we know that $\frac{dm}{dt}$ will be $50\%$ of the current mass (by the same logic that we know that $\frac{dm}{dt}$ of $e^t$, which is the function representing $100\%$ growth, is $100\%$ of the current mass). More generally
$$\frac{dm}{dt} = \frac{d}{dt}(e^{g(t)}) = rm$$
where $r$ is the growth rate ($1$ for $100\%$, $0.5$ for $50\%$, etc.), and $m$ is mass. We also know, by the chain rule, that
$$\frac{dm}{dt} = e^{g(t)} \cdot g'(t)$$
and therefore
$$rm = e^{g(t)} \cdot g'(t)$$
Now, since $m = e^{g(t)}$
$$rm = m * g'(t)$$
$$r = g'(t)$$
Now we want to find out what $g(t)$ is, so we can find out what $x$ is (remember, we're modeling $e^x$ as $e^{g(t)}$). If we integrate both sides we get
$$g(t) = rt + C$$
So $e^x$ is $e^{rt+c}$, which is equal to $e^{rt} \cdot e^C$.
$e^C$ is just another constant, $C$, and, as shown in johnnyb's answer, $C$ is the starting mass, which we choose to be $1$ for this example.
So, we are left with $$m = e^{rt}$$
This is a very cool result: It shows that we can model continuous growth (where all the tissue is active!) for any desired growth rate, and for any time, using $e^{rt}$. With $e$ as a base, the overall rate (the product of the growth rate and duration) is represented by the exponent.
Now we are ready to answer your question:
We can actually model the noncontinuous scenario (red line) using a continuous function, $2^t$ (blue line)
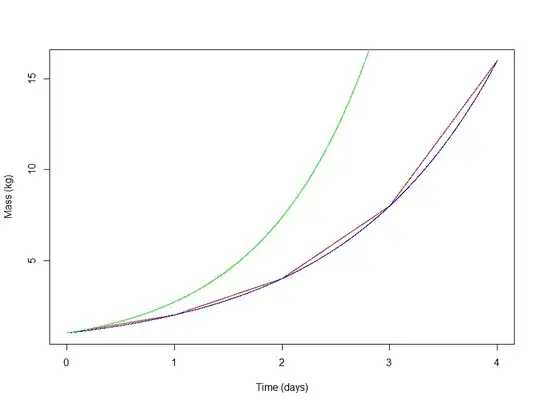
This is certainly a continuous compounding scenario, where all the tissue is active, but the growth rate is clearly less than $100\%$, since it rises slower than $e^t$ (green line). So, what is the growth rate of $2^t$? Well we can rewrite $2^t$ as follows
$$2^t = b^{\log_b (2^t)}$$ where $b$ is any arbitrary base. Let's see what happens when we pick $e$ as our base.
$$2^t = e^{ln(2^t)}$$
Now, given that $ln{(2^t)} = t \cdot ln(2)$, we have
$$2^t = e^{ln(2)\cdot t}$$
which means that the growth rate of $2^t$ is $ln(2)$.
Now we see why it is convenient to model population growth with base $e$ , even if we don't care about finding the time derivative. With base $e$, the rate is encoded directly in the exponent. It's explicit! With another base, $b$, we need to multiply the exponent by a constant, $ln(b)$, to calculate the rate.
Using $e$ here serves two purposes - it makes it easier to infer the growth rate of whatever function is being described. When we see $e^{rt}$, we know right away that the growth rate is $r$. And remember, the growth rate, $r$, is fundamental in describing the system - it is an invariant. Secondly, if we are trying to model a physical phenomenon, and we have actually empirically measured the growth rate or theoretically inferred it, we can readily model the phenomenon using $e^{rt}$, or indeed $A \cdot e^{rt}$.
Remember, all exponential curves belong to the same family, and any given curve can be expressed using any given base. Using base $e$ is simply a very convenient base.